Email Alert | RSS 帮助

中国防痨杂志 ›› 2020, Vol. 42 ›› Issue (2): 133-142.doi: 10.3969/j.issn.1000-6621.2020.02.010
张蓝月,耿艺漫,贾红彦,肖婧,李自慧,潘丽萍,孙义成( ),张宗德()
),张宗德()
ZHANG Lan-yue,GENG Yi-man,JIA Hong-yan,XIAO Jing,LI Zi-hui,PAN Li-ping,SUN Yi-cheng(),ZHANG Zong-de()
摘要:
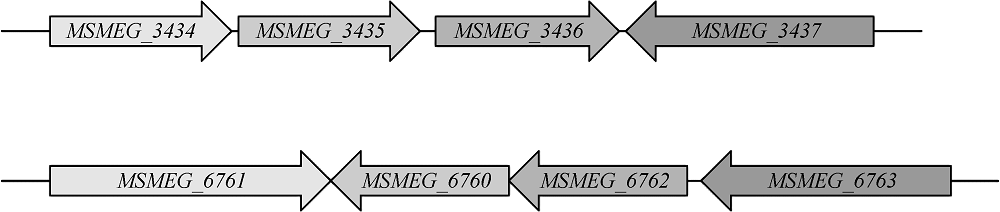
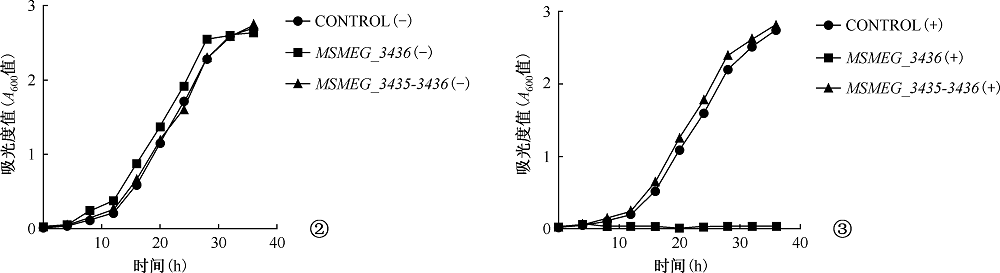
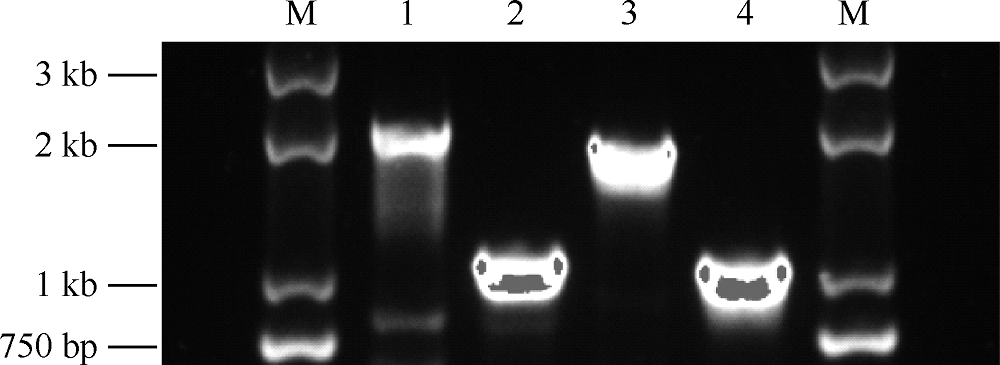
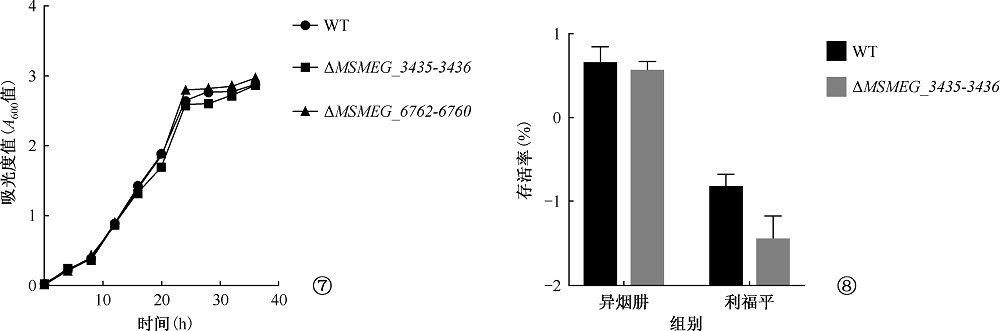
目的 初步探讨耻垢分枝杆菌(Mycobacterium smegmatis)毒素-抗毒素(toxin-antitoxin,TA)系统基因的功能及其在细菌药物耐受中的作用。方法 利用无水四环素(ATc)诱导穿梭质粒构建毒素基因(MSMEG_3436和MSMEG_6760)表达系统,检测毒素基因表达的抑菌作用。应用CRISPR-Cas12a基因编辑技术构建ΔMSMEG_3435-3436和ΔMSMEG_6762-6760敲除菌株,探究毒素-抗毒素系统对菌株生长的影响。通过计算菌株存活率检测MSMEG_3435-3436基因对异烟肼(96μg/ml)和利福平(40μg/ml)的耐受相关性。在耻垢分枝杆菌中用LacZ报告基因分别替换毒素-抗毒素基因(MSMEG_1277-1278、 MSMEG_1283-1284、 MSMEG_3435-3436、 MSMEG_4447-4448和MSMEG_5635-5634),构建5个启动子活性检测突变菌株(SY3328、SY3309、SY6407、SY3310和SY3311),并将pMV261空载体和pMV261-抗毒素系列质粒分别电转至5个突变菌株中,通过测定吸光度值(A600、A550和A420)计算β-半乳糖苷酶活性[酶活性单位为“Miller单位(MU)”],以检测毒素抗毒素系统的启动子活性。结果 在耻垢分枝杆菌中,ATc诱导表达毒素基因MSMEG_3436可抑制细菌生长,而同时表达对应的抗毒素基因MSMEG_3435可消除抑制作用;ATc诱导表达毒素基因MSMEG_6760未发现明显的抑菌作用。与野生株相比,ΔMSMEG_3435-3436和ΔMSMEG_6762-6760敲除菌株在7H9液体培养基中生长表型无明显差异。野生株和ΔMSMEG_3435-3436敲除菌株经异烟肼和利福平处理后的存活率[分别为(4.38±1.48)%和(3.49±0.66)%,(0.15±0.04)%和(0.03±0.02)%]显示毒素-抗毒素基因MSMEG_3435-3436与药物耐受性无关(t=0.548,P=0.613;t=2.663,P=0.056)。启动子(SY3328、SY3309、SY6407、SY3310和SY3311)在携带pMV261-空载体和pMV261抗毒素表达质粒的报告菌株中的β-半乳糖苷酶活性分别为(376.50±17.13)和(315.50±20.71)、(189.00±12.24)和(160.70±9.89)、(225.20±9.95)和(211.70±2.57)、(221.40±12.07)和(186.60±13.17)、(179.10±5.87)和(127.70±19.21)MU,差异均无统计学意义(t=2.272,P=0.086;t=1.795,P=0.147;t=1.319,P=0.258;t=1.949,P=0.123;t=2.562,P=0.063)。结论 成功构建了MSMEG_3435-3436和MSMEG_6762-6760在耻垢分枝杆菌中的诱导表达体系及敲除菌株,并发现MSMEG_3435-3436是一个新的有功能的毒素-抗毒素系统,以及这两个毒素-抗毒素系统与菌株生长表型及异烟肼和利福平耐受性无关,最后发现耻垢分枝杆菌中5对毒素-抗毒素系统的抗毒素基因可能在自身启动子调控中不发挥关键作用,可为进一步研究结核分枝杆菌毒素-抗毒素系统的功能提供线索。
 京公网安备11010202007215号
ip访问总数: ip当日访问总数: 当前在线人数:
京公网安备11010202007215号
ip访问总数: ip当日访问总数: 当前在线人数:
 本作品遵循Creative Commons Attribution 3.0 License授权许可
本作品遵循Creative Commons Attribution 3.0 License授权许可